Concentración y dilución de orina.
CONCENTRACION Y DILUCION DE ORINA; REGULACION DE LA OSMOLARIDAD DEL LIQUIDO EXTRACELULAR Y DE LA CONCENTRACION DE SODIO
Para el correcto funcionamiento de las células del organismo, estas deben estar bañadas en líquido extracelular con una concentración relativamente constante de electrólitos y otros solutos. La concentración total de solutos en el líquido extracelular (y, por tanto, la osmolaridad) está determinada por la cantidad de soluto dividida por el volumen de líquido extracelular. De este modo, la concentración de sodio y la osmolaridad del líquido extracelular están, en gran parte, reguladas por la cantidad de agua extracelular.
El agua corporal total está controlada por: 1) la ingestión de líquido, que está regulado por los factores que determinan la sed, y 2) por la excreción renal de agua, controlada por los múltiples factores que influyen en la filtración glomerular y la reabsorción tubular.

La hormona antidiurética controla la concentración de la orina
Existe un sistema de retroalimentación potente para regular la osmolaridad y la concentración de sodio en el plasma que actúa modificando la excreción renal de agua con independencia de la excreción de solutos. Un efector fundamental de esta retroalimentación es la hormona antidiurética (ADH), también llamada vasopresina.
Los riñones conservan agua excretando una orina concentrada
La capacidad del riñón de formar una orina que está más concentrada que el plasma es esencial para la supervivencia de los mamíferos que viven en la tierra, incluidos los seres humanos. El agua se pierde continuamente a través de diversas vías, como los pulmones por evaporación hacia el aire espirado, el aparato digestivo a través de las heces, la piel a través de la evaporación y la sudoración y los riñones a través de la excreción de orina. Es necesario ingerir líquido para cubrir esta pérdida, pero la capacidad del riñón de formar un volumen pequeño de orina concentrada minimiza la ingestión de líquido necesaria para mantener la homeostasis, una función que es especialmente importante cuando escasea el agua.
Requisitos para excretar una orina concentrada: concentraciones altas de ADH y médula renal hiperosmótica

Función del tùbulo distal y de los conductos colectores en la excreción de una orina concentrada
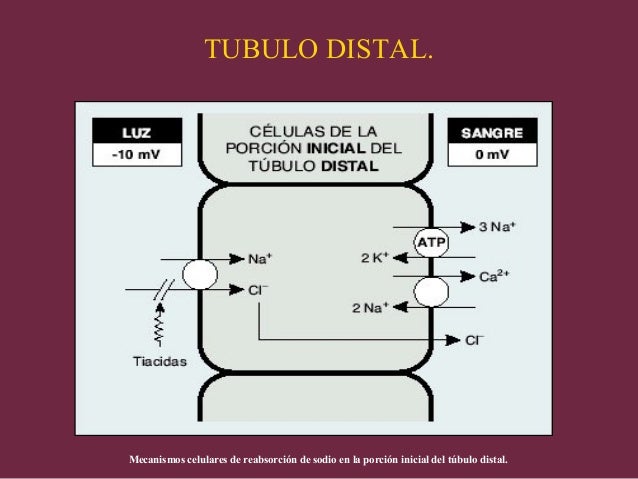
La primera parte del tubulo distai diluye más el líquido tubular porque este segmento, como el asa ascendente de Henle, transporta de forma activa cloruro de sodio fuera del tùbulo, pero es relativamente impermeable al agua. A medida que el líquido fluye hacia el tùbulo colector cortical, la cantidad de agua reabsorbida depende mucho de la concentración plasmática de ADH.
Si falta la ADH, este segmento es casi impermeable al agua y no reabsorbe agua sino que continúa reabsorbiendo solutos y diluye más la orina. Cuando hay una concentración alta de ADH, el tùbulo colector cortical se hace muy permeable al agua, de manera que se reabsorben ahora grandes cantidades de agua desde el tùbulo hacia el intersticio de la corteza, donde es barrida por el flujo rápido de los capilares peritubulares. El hecho de que estas grandes cantidades de agua se reabsorban hacia la corteza, en lugar de hacia la médula renal, ayuda a conservar la elevada osmolaridad del líquido intersticial medular.
Si falta la ADH, este segmento es casi impermeable al agua y no reabsorbe agua sino que continúa reabsorbiendo solutos y diluye más la orina. Cuando hay una concentración alta de ADH, el tùbulo colector cortical se hace muy permeable al agua, de manera que se reabsorben ahora grandes cantidades de agua desde el tùbulo hacia el intersticio de la corteza, donde es barrida por el flujo rápido de los capilares peritubulares. El hecho de que estas grandes cantidades de agua se reabsorban hacia la corteza, en lugar de hacia la médula renal, ayuda a conservar la elevada osmolaridad del líquido intersticial medular.
El intercambio por contracorriente en los vasos rectos conserva la hiperosmolaridad en la médula renal
A la médula renal debe llegar un flujo de sangre que cubra las necesidades metabólicas de las células de esta parte del riñón. Sin un sistema de flujo sanguíneo medular especial los solutos bombeados a la médula renal por el sistema multiplicador por contracorriente se disiparían rápidamente. El flujo sanguíneo de la médula renal tiene dos características que contribuyen a conservar las elevadas concentraciones de solutos:1. El flujo sanguíneo medular es bajo, suponiendo menos de un 5% del flujo sanguíneo renal tota! Este flujo sanguíneo lento es suficiente para cubrir las necesidades metabólicas de los tejidos, pero ayuda a minimizar la pérdida de solutos del intersticio medular.
2. Los vasos rectos sirven de intercambiadores por contracorriente, lo que minimiza el lavado de solutos del intersticio medular.
El aumento del flujo sanguíneo medular reduce la capacidad de concentrar la orina.
Ciertos vasodilatadores pueden aumentar de forma acentuada el flujo sanguíneo en la médula renal, con lo que «lavan» parte de los solutos de la médula renal y reducen la capacidad máxima de concentrar la orina. Incrementos grandes de la presión arterial pueden aumentar también el flujo sanguíneo de la médula renal en mayor grado que en otras regiones del riñón y tender a lavar el intersticio hiperosmótico, lo que reduce la capacidad de concentración de la orina.Resumen del mecanismo de concentración de la orina y de los cambios en la osmolaridad en diferentes segmentos de los túbulos

Túbulo proximal: Alrededor del 65% de los electrólitos filtrados se reabsorben en el túbulo proximal. Pero las membranas tubulares proximales son muy permeables al agua, de manera que siempre que se reabsorben solutos, el agua también difunde a través de la membrana tubular por osmosis.
Asa descendente de Henle: A medida que el líquido fluye por el asa descendente de Henle, el agua se reabsorbe hacia la médula. La rama descendente es muy permeable al agua, pero mucho menos al cloruro de sodio y a la urea.
Asa ascendente fina de Henle: La rama ascendente fina es prácticamente impermeable al agua, pero reabsorbe parte del cloruro de sodio. Debido a la elevada concentración del cloruro de sodio en el líquido tubular, y por la extracción de agua del asa descendente de Henle, hay una difusión pasiva del cloruro de sodio desde la rama ascendente fina hacia el intersticio medular.
Asa ascendente gruesa de Henle. La parte gruesa del asa ascendente de Henle es prácticamente impermeable al agua, pero grandes cantidades de sodio, cloro y potasio y otros tipos de iones se transportan activamente desde el tùbulo hacia el intersticio medular.
Primera parte del tùbulo distal: La primera parte del tùbulo distai tiene propiedades similares a las del asa ascendente gruesa de Henle, de manera que la dilución del líquido tubular a unos 50 mOsm/1 tiene lugar a medida que los solutos se reabsorben mientras el agua permanece en el tùbulo.
Parte final del tùbulo distai y túbulos colectores corticales: Al final del tùbulo distai y en los túbulos colectores corticales, la osmolaridad del líquido depende de la concentración de ADH. Con concentraciones altas de ADH, estos túbulos son muy permeables al agua, y se reabsorben cantidades significativas de agua. Pero la urea no es muy difusible en esta parte de la nefrona, lo que da lugar a una mayor concentración de la urea a medida que se reabsorbe el agua. Esto permite que la mayor parte de la urea que llega al tùbulo distai y al conducto colector pase a los conductos colectores medulares internos, desde donde al final se reabsorbe o excreta en la orina.
Conductos colectores medulares internos: La concentración de líquido en los conductos colectores medulares internos también depende de: 1) la ADH y 2) la osmolaridad del intersticio medular circundante establecida por el mecanismo de contracorriente.
Centros de la sed en el sistema nervioso central
Las neuronas del centro de la sed responden a las inyecciones de soluciones hipertónicas de sal estimulando la búsqueda de agua. Estas células funcionan casi con toda seguridad como osmorreceptores para activar el mecanismo de la sed, de la misma forma que los osmorreceptores estimulan la liberación de ADH. El aumento de la osmolaridad del líquido cefalorraquídeo en el tercer ventrículo tiene prácticamente el mismo efecto favorecedor de la búsqueda de agua.
Estímulos de la sed
Algunos de los estímulos conocidos de la sed. Uno de los más importantes es el aumento de la osmolaridad del líquido extracelular, que provoca una deshidra- tación intracelular en los centros de la sed, lo que estimula la sensación de sed. El valor de esta respuesta es obvia: ayuda a diluir los líquidos extracelulares y normaliza la osmolaridad. Las reducciones del volumen del líquido extracelular y de la presión arterial también estimulan la sed a través de una vía que es independiente de la estimulada por la osmolaridad plasmática. Luego la pérdida de volumen sanguíneo por una hemorragia estimula la sed incluso aunque no cambie la osmolaridad plasmática. Esto se debe probablemente a impulsos neurales procedentes de los barorreceptores car- diopulmonares y arteriales sistémicos en la circulación. Un tercer estímulo importante de la sed es la angiotensina II. Los estudios en animales han mostrado que la angiotensina II actúa sobre el órgano subfornical y sobre el órgano vasculoso de la lámina terminal la estimulan factores asociados a lahipovolemia y la presión arterial baja, su efecto sobre la sed ayuda a restaurar el volumen sanguíneo y la presión arterial hacia valores normales, junto a las otras acciones de la angiotensina II sobre los riñones para reducir la excreción de líquido.

Comentarios
Publicar un comentario